Chiroptera
Bats
Nancy B. Simmons and Tenley Conway


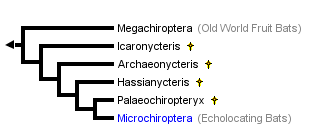

This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The clade Chiroptera includes two extant clades, Megachiroptera (Old World Fruit Bats) and Microchiroptera (echolocating bats). In addition, Chiroptera includes at least four extinct clades that are most closely related to Microchiroptera. There are over nine hundred extant species of bats (Koopman, 1993). Bats vary greatly in size. The smallest bat, Craseonycteris thonglongyai (Microchiroptera), weighs less than 2 g and has a wingspan of 12-13 cm, while the largest bats, those of the genus Pteropus (Megachiroptera), weigh up to 1.5 kg and may have a wing span over 2m (Fenton, 1992).
Bats are unique among mammals as they are the only group to have evolved true powered flight. Some other mammals such as "flying" squirrels and "flying" lemurs can glide through the air for long distances, but they are not capable of sustained flight. In contrast, bats can propel themselves with their wings, gaining and loosing altitude and flying for long periods.
Bats are nocturnal and usually spend the daylight hours roosting in caves, rock crevices, trees, or manmade structures such as houses and/or bridges. Some bats are solitary, while others are found in colonies that may include over a million individuals.
Activity begins around dusk, when bats leave the day roost and start feeding. The clade Chiroptera includes species with very diverse food preferences, including bats that eat either meat, insects, fish, fruit, nectar, or a variety of food types. Only three species of bats actually feed on blood Desmodontinae). Many bats remain at their feeding sites until just before dawn when they return to the day roost.
Classification outlines the higher-level classifications within Chiroptera.
Characteristics
Unique Characteristics
One unique feature of bats is their modified forelimbs, which support a wing membrane (patagium). The basic elements of the mammalian limb are present in bats, although the relative sizes of most bones and muscles differ from those of nonflying mammals. The most elongated parts of the limb are those of the hand (metacarpal bones) and fingers (phalanges). The primary functions of these bones in bats is to provide support for the patagium and control its movements. The patagium stretches between the fingers and attaches to the side or back of the bat and the lower leg. Part of the membrane extends between the hindlimbs. Numerous blood vessels and nerves are present throughout the wing membrane. Bats also have five unique muscles present in the patagium, and use additional muscles in the chest and back to move the wings up and down.
The most obvious difference between bird wings and those of bats is that bird wings are made of feathers, not a skin membrane. Birds have an elongated arm, but do not have elongated fingers like bats. Additionally, the muscles used in both the upstroke and downstroke are found in the chest of birds, while the upstroke muscles are on the back in bats (Fenton, 1983).
The orientation of the hindlimb is also unique to bats. The hip joint is rotated 90° so that the legs project sideways and the knee faces almost backwards. Due in part to the rotation of the hindlimb, the walking motion of bats differs from other tetrapods, often appearing awkward. The hindlimb is designed to support the patagium in flight and allow the bat to roost hanging from its hindlimbs. Most bats have a tendon system in the toes that locks the claws in place so the bat can hang upside down even when asleep.
Bats have other unique characteristics including many morphological synapomorphies.
General Characteristics
The body of a bat is ventrally compressed with a short neck region. The bones tend to be slender and light-weight. The majority of the body weight is concentrated in the chest region due to the large flight muscles.
The overall shape of the head varies more in bats than within most other groups of mammals. Some bats have very elongated muzzles while others have broad, short faces. There is a correlation between the shape of the head and the type of food eaten. For example, most nectar feeders have long, narrow muzzles that are good for reaching into flowers, while many fruit eaters have short, broad faces good for biting rounded fruits (Hill and Smith, 1984).
The ears range from small and round to large and pointed, and often have a cartilaginous fold (tragus) present at the notch of the ear. There is additional variation in the nasal and lip regions of bats. Some bats have complex noseleafs, folds, or wrinkles on their muzzles. The function of facial ornamentation is not well understood, although it may effect the emission of echolocation calls in some taxa (Fenton, 1992).
A major misconception about bats is that they are blind. This idea originated from the fact that bats are able to successfully maneuver in the dark and often have small eyes. While some bats do have very small eyes (most Microchiroptera) many have large and complex eyes (Megachiroptera). Experiments on several species of bats have shown that they are able to distinguish patterns even at low light levels (Hill and Smith, 1984).
Bats usually have black or brown fur, although the fur can also be gray, white, red, or orange. In some species there are stripes on the face or down the back, or patches of white on the face or above the shoulder. The length of the fur also varies among species from short and dense to long and fluffy. The wing membrane is usually dark in color, although it may have white on the tips or be a lighter color around the bones in the membrane. A few bats have white or pale yellow wings. There are also little hairs on the membrane itself. These hairs can be the color of the wing or the same color as the body.
Bat Monophyly
Until the 1970s, most evolutionary biologists assumed that bats form a monophyletic group. Recently, however, several authors have questioned monophyly of Chiroptera (Jones and Genoways, 1970; Smith, 1976, 1980; Smith and Madkour, 1980; Hill and Smith, 1984; Pettigrew, 1986, 1991a, 1991b; Pettigrew and Jamieson, 1987; Pettigrew et al., 1989) creating what has become known as the “bat monophyly controversy”. Proponents of the hypothesis that bats are diphyletic pointed out that many similarities between Megachiroptera and Microchiroptera involve the flight mechanism. It is therefore possible that convergent evolution of aerial locomotion, rather than shared ancestry, might account for the similarities found between megachiropteran and microchiropteran bats (Jones and Genoways, 1970).
The bat monophyly hypothesis states the Megachiroptera and Microchiroptera are each others closest relatives in an evolutionary sense (i.e., they form a clade). If this is true, then their shared characteristics, including the ability to fly, would have been present in their most recent common ancestor (Simmons, 1994; 1995). It follows from this that there was only one origin of powered flight in mammals. In contrast, the diphyly hypothesis states that megachiropteran and microchiropteran bats do not form a monophyletic group, instead having evolved independently from two different groups of non-flying mammals. It has been suggested that Megachiroptera is more closely related to Dermoptera and Primates than to Microchiroptera (Smith and Madkour, 1980; Pettigrew, 1986, 1991a, b, 1995; Pettigrew and Jamieson, 1987; Pettigrew et. al., 1989). In this case, the characteristics common to both groups of bats either evolved as a result of convergent evolution or are simply the result of retention of primitive features. If bats are diphyletic, the ability to fly must have evolved once in Megachiroptera and again in Microchiroptera.
There have been numerous studies using biochemical, molecular, and/or morphological data to analyze the relationship between Megachiroptera, Microchiroptera and other taxa. Diphyly has only been supported in two data sets: features of the nervous system (Pettigrew, 1986, 1991a, 1991b; Pettigrew et. al., 1989; Johnson and Kirsch, 1993) and of the penis (Smith and Madkour, 1980). In contrast, monophyly has been supported in studies examining a large and diverse set of morphological features, including those of the nervous and reproductive systems (Luckett, 1980a, 1993; Wible and Novacek, 1988; Kovtun, 1989; Thewissen and Babcock, 1991, 1993; Kay et. al., 1992; Novacek, 1992, 1994; Beard, 1993; Simmons, 1993a, 1994, 1995; Wible and Martin, 1993; Simmons and Quinn, 1994, Miyamoto, 1996), DNA-DNA hybridization data (Kirsch et al., 1995; Hutcheon and Kirsch, 1996; Kirsch, 1996), and DNA nucleotide sequence data from mitochondrial and nuclear genes (Adkins and Honeycutt, 1991, 1993, 1994; Mindell et al., 1991; Ammerman and Hillis, 1992; Bailey at al., 1992; Stanhope et al., 1992, 1993, 1996; Honeycutt and Adkins, 1993; Knight and Mindell, 1993; Novacek, 1994, Allard et al., 1996; Miyamoto, 1996; Porter et al., 1996).
Because the vast majority of available data strongly support a sister-group relationship between Megachiroptera and Microchiroptera, bat monophyly is now regarded as a very strongly supported hypothesis.
Discussion of Phylogenetic Relationships
The fossil record of bats extends back at least to the early Eocene, and chiropteran fossils are known from all continents except Antarctica. Icaronycteris, Archaeonycteris, Hassianycteris, and Palaeochiropteryx, unlike most other fossil bats, have not been referred to any extant family or superfamily. These Eocene taxa are known from exceptionally well-preserved fossils, and they have long formed a basis for reconstructing the early evolutionary history of Chiroptera (see review in Simmons and Geisler, 1998).
Smith (1977) suggested that these taxa represent an extinct clade of early microchiropterans ("Palaeochiropterygoidea"). In contrast, Van Valen (1979) argued that these fossil forms are representatives of a primitive grade ancestral to both Megachiroptera and Microchiroptera ("Eochiroptera"). Novacek (1987) reanalyzed morphology of Icaronycteris and Palaeochiropterx and concluded that they are more closely related to Microchroptera than to Megachiroptera. Most recently, Simmons and Geisler (1998) found that Icaronycteris, Archaeonycteris, Hassianycteris, and Palaeochiropteryx represent a series of consecutive sister-taxa to extant microchiropteran bats.
Geographic Distribution
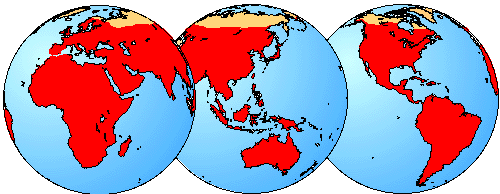
The geographical distribution of bats is shown in red. Distribution from Hill and Smith (1984).
References
Adkins, R. M., & R. L. Honeycutt. 1991. Molecular phylogeny of the superorder Archonta. Proceedings of the National Academy of Science U.S.A. 88: 10317-10321.
Adkins, R. M., & R. L. Honeycutt. 1993. A molecular examination of archontan and chiropteran monophyly. In R. D. E. MacPhee (ed.), Primates and their relatives in phylogenetic perspective, pp. 227-249. Advances in Primatology Series. New York: Plenum.
Adkins, R. M., & R. L. Honeycutt. 1994. Evolution of the primate cytochrome oxidase subunit II gene. Journal of Molecular Evolution 38: 215-231.
Allard, M. W., B. E. McNiff, & M. M. Miyamoto. 1996. Support for interordinal eutherian relationships, with an emphasis on Primates and their archontan relatives. Molecular Phylogenetic Evolution 5: 78-88.
Ammerman, L. K., & D. M. Hillis. 1992. A molecular test of bat relationships: monophyly or diphyly? Systmatic Biology 41: 222-232.
Altringham, J. D. 1996. Bats : Biology and Behavior. New York: Oxford University Press.
Bailey, W. J., Slightom, J. L., & Goodman, M. 1992. Rejection of the “flying primate” hypothesis by phylogenetic evidence from the e-globin gene. Science 256: 86-89.
Barbour, R .W., & W. H. Davis. 1969. Bats of America. Lexington: University Press of Kentucky.
Beard, K. C. 1993. Phylogenetic systematics of Primatomorpha, with special reference to Dermoptera. In F. S. Szalay, M. J. Novacek, and M. C. McKenna (eds.), Mammal phylogeny: Placentals, pp. 129-150. New York: Springer-Verlag.
Fenton, M. B. 1983. Just Bats. Toronto: University of Toronto Press.
Fenton, M. B. 1992. Bats. New York: Facts On File.
Findley, J. S. 1993. Bats: a community perspective. Cambridge: Cambridge University Press.
Hill, J. E. & J. D. Smith. 1984. Bats: a natural history. Austin: University of Texas Press.
Honeycutt, R. L., & R. M. Adkins. 1993. Higher-level systematics of eutherian mammals: an assessment of molecular characters and phylogenetic hypotheses. Annual Revue of Ecology and Systematics 24: 27-306.
Hutcheon, J. M., & J. A. W. Kirsch. 1996. Inter-familial relationships within the Microchiroptera: a preliminary study using DNA hyrbridization. Bat Research News 36: 73-74.
Johnson, J. I., & J. A. W. Kirsch. 1993. Phylogeny through brain traits: interordinal relationships among mammals including Primates and Chiroptera. In R. D. E. MacPhee (ed.), Primates and their relatives in phylogenetic perspective, pp. 293-331. Advances in Primatology Series. New York: Plenum.
Jones, J. K., & H. H. Genoway. 1970. Chiroptera systematics. In R, H, Slaughter and D.W. Walton (eds.), About bats: a chiropteran symposium, pp. 3-21. Dallas, TX: Southern Methodist Univ. Press.
Kay, R. F., J. G. M. Thewissen, & A. D. Yoder. 1992. Cranial anatomy of Ignacius graybullianus and the affinities of the Plesiadapiformes. American Journal of Physical Anthropology 89: 477-498.
Kirsch, J. A. W. 1996. Bats are monophyletic; megabats are monophyletic; but are microbats also? Bat Research News 36: 78.
Kirsch, J. A. W., T. F. Flannery, M. S. Springer, & F.-J. Lapointe. 1995. Phylogeny of the Pteropodidae (Mammalia: Chiroptera) based on DNA hybridization, with evidence for bat monophyly. Australian Journal of Zoology 43: 395-428.
Kirsch, J. A., J. M. Hutsceon, D. G. Byrnes & B. D. Lloyd. 1998. Affinites and hidtorical zoogeography of the New Zealand Short-tailed bat, Mystacina tuberculata Gray 1843, inferred from DNA-hybridization comparisons. Journal of Mammalian Evolution 5(1): 33-64.
Knight, A., and D. P. Mindell. 1993.Substitution bias, weighting of DNA sequence evolution, and the phylogenetic position of Fea’s viper. Systamatic Zoolology 38: 7-25.
Koopman, K. F. 1993. Chiroptera. In Mammal Species of the World. D.E. Wilson and D.M. Reeder (eds.) Washington: Smithsonian Institution Press.
Kovtun, M. F. 1989. On the origin and evolution of bats. In V. Hanák, I. Horácek, and J. Gaisler (eds.), European bat research 1987, pp. 5--12. Praha: Charles Univ. Press.
Kunz, T. H. 1982. Ecology of Bats. New York: Plenum Press.
Luckett, W. P. 1980. The use of fetal membrane characters in assessing chiropteran phylogeny. In D. E. Wilson and A. L. Gardner (eds.), Proceedings Fifth international bat research conference, pp. 245-266. Lubbock: Texas Tech Press.
Luckett, W. P. 1993. Developmental evidence from the fetal membranes for assessing archontan relationship. In R. D. E. MacPhee (ed.), Primates and their relatives in phylogenetic perspective. Adv. Primatol. Ser., pp. 149 186. New York: Plenum.
Mindell, D. P., C. W. Dick, & R. J. Baker. 1991. Phylogenetic relationships among megabats, microbats, and primates. Proc. Natl. Acad. Sci. U.S.A. 88: 10,322-10,326.
Miyamoto, M. M. 1996. A congruence study of molecular and morphological data for eutherian mammals. Mol. Phylogenet. Evol. 6: 373-390.
Miyamoto, M. M. 1992. Fossils as critical data for phylogeny. In M. J. Novacek and Q. D. Wheeler (eds.), Extinction and phylogeny, pp. 46-88. New York: Columbia Univ. Press.
Miyamoto, M. M. 1994. Morphological and molecular inroads to phylogeny. In L. Grande and O. Rieppel (eds), Interpreting the hierarchy of nature: from systematic patterns to evolutionary process theories, pp. 85-131. New York: Academic Press.
Pettigrew, J. D. 1986. Flying primates? Megabats have the advanced pathway from eye to midbrain. Science 231: 1304-1306.
Pettigrew, J. D. 1988. Microbat vision and echolocation in an evolutionary context. In P. E. Nachtigill and P. W. B. Moore (eds.), Animal sonar: processes and performance, pp. 645-650. New York: Plenum.
Pettigrew, J. D. 1991a. Wings or brain? Convergent evolution in the origins of batc. Systamatic Zoology 40: 199-216.
Pettigrew, J. D. 1991b. A fruitful, wrong hypothesis? Response to Baker, Novacek, and Simmons. Systamatic Zoology 40: 231-239.
Pettigrew, J. D. 1995. Flying primates: crashed, or crashed through? In P. A. Racey and S. M. Swift (eds.), Ecology, evolution and behavior of bats. Symposium of the Zoological Society of London 67: 3-26.
Pettigrew, J. D., & B. G. M. Jamieson. 1987. Are flying foxes (Chiroptera: Pteropodidae) really primates? Australian Mammal. 10: 119-124.
Pettigrew, J. D., B. G. M. Jamieson, S. K. Robson, L. S. Hall, K. I. McAnally, & H. M. Cooper. 1989. Phylogenetic relations between microbats, megabats and primates (Mammalia: Chiroptera and Primates). Philosophical Transactions of the Royal Society of London B 325: 489-559.
Porter, C. A., M. Goodman, & M. J. Stanhope. 1996. Evidence on mammalian phylogeny from sequences of exon 28 of the von Willebrand Factor gene. Molecular Phylogenetic Evolution 5: 89-101.
Richarz, K. & A. Limbrunner. 1993. The World of Bats. Neptune City, NJ: T.F.H. Publications, Inc.
Ruttle, M. D. 1988. America's Neighborhood Bats. Austin: University of Texas Press.
Simmons, N. B. 1993. The importance of methods: archontan phylogeny and cladistic analysis of morphological data. In R. D. E. MacPhee (ed.), Primates and their relatives in phylogenetic perspective, pp. 1--61. New York: Plenum.
Simmons, N. B. 1994. The case for chiropteran monophyly. American Museum Novitates 3103: 54 pp.
Simmons, N. B. 1995. Bat relationships and the origin of flight. In P. A. Racey and S. M. Swift (eds.), Ecology, evolution and behavior of bats. Symposium of the Zoological Society of London 67: 27-43.
Simmons, N. B. 1998. A reappraisal of interfamilial relationships of bats. In Bats: Phylogeny, Morphology, Echolocation, and Conservation Biology. T. H. Kunz and P.A. Racey (eds.) Washington: Smithsonian Institution Press
Simmons, N. B. & J.H. Geisler. 1998. Phylogenetic relationships of Icaronycteris, Archeonycteris, Hassianycteris, and Palaeochiropteryx to extant bat lineages, with comments on the evolution of echolocation and foraging strategies in microchiroptera. Bulletin of the American Museum of Natural History. 235:1-182.
Simmons, N. B., & T. H. Quinn. 1994. Evolution of the digital tendon locking mechanism in bats and dermopterans: a phylogenetic perspective. Journal of Mammalian Evolution 2: 231-254.
Smith, J. D. 1976. Chiropteran evolution. In R. J. Baker, J. K. Jones, and D. C. Carter (eds.), Biology of bats of the New World family Phyllostomatidae, part I, pp. 49-69. Special Publ. The Museum, Texas Tech Univ., vol. 10. Lubbock: Texas Tech Univ.
Smith, J. D. 1980. Chiropteran phylogenetics: introduction. In D. E. Wilson and A. L. Gardner (eds.), Proceedings Fifth International Bat Research Conference, pp. 233-244. Lubbock: Texas Tech Press.
Smith, J. D., & G. Madkour. 1980. Penial morphology and the question of chiropteran phylogeny. In D. E. Wilson and A. L. Gardner (eds.), Proceedings Fifth International Bat Research Conference, pp. 347-365. Lubbock: Texas Tech Press.
Stanhope, M. J., W. J. Bailey, J. Czelusniak, M. Goodman, J.-S. Si, J. Nickerson, J. G. Sgouros, G. A. M. Singer, & T. K. Kleinschimidt. 1993. A molecular view of primate supraordinal relationships from the analysis of both nucleotide and amino acid sequences. In R. D. E. MacPhee (ed.), Primates and their relatives in phylogenetic perspective, pp. 251-292. Adv. Primatol. Ser. New York: Plenum.
Stanhope, M. J., J. Czelusniak, J.-S. Si, J. Nickerson, & M. Goodman. 1992. A molecular perspective on mammalian evolution from the gene encoding interphotoreceptor retinoid binding protein, with convincing evidence for bat monophyly. Molecular Phylogenetic Evolution 1: 148-160.
Stanhope, M. J., M. R. Smith, V. G. Waddell, C. A. Porter, M. S. Thewissen, J. G. M., & S. K. Babcock. 1991. Distinctive cranial and cervical innervation of wing muscles: new evidence for bat monophyly. Science 251: 934-936.
Thewissen, J. G. M., & S. K. Babcock. 1993. The implications of the propatagial muscles of flying and gliding mammals for archontan systematics. In R. D. E. MacPhee (ed.), Primates and their relatives in phylogenetic perspective, pp. 91-109. Advanced Primatology Series. New York: Plenum.
Wible, J. R., & J. R. Martin. 1993.Ontogeny of the typmanic floor and roof in archontans. In R. D. E. MacPhee (ed.), Primates and their relatives in phylogenetic perspective, pp. 111-148. Advanced Primatology Series. New York: Plenum.
Wible, J. R., & M. J. Novacek. 1988. Cranial evidence for the monophyletic origin of bats. American Museum Novitates 2911: 19 pp.
Wilson, D. E. 1997. Bats in Question. Washington: Smithsonian Institution Press.
Information on the Internet
For more information on Chiroptera, see these web sites:
- Bats Conservation International. Includes bat trivia, photographs, conservation news and information on bat houses.
- University of California Museum of Paleontology. Information on ecology, systematics, morphology and the fossil record of bats.
- James Buzbee's Bat House. Hundreds of links to pages about bats including regional sites, exhibits, organizations, sites for kids and pictures.
- Incredible Bats. Dispells common myths about bats.
- Bat Research Laboratory- at Purdue University. A research project trying to develop a bio-sonar model for target recognition and classification based on a bats echolocation system.
- Pacific Northwest Bat Call Library. A collection of bat calls and links to other bat call libraries.
Title Illustrations
Molossus rufus (Microchiroptera) and Phylostomus hastatus (Microchiroptera)

| Scientific Name | Molossus rufus |
|---|---|
| Image Use |
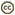 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 1998 Nancy B. Simmons

|

| Scientific Name | Phylostomus hastatus |
|---|---|
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 1994 Nancy B. Simmons
|
About This Page
Nancy B. Simmons
American Musuem of Natural History, New York, New York, USA
Tenley Conway
University of Toronto at Mississauga, Ontario, Canada
Correspondence regarding this page should be directed to Nancy B. Simmons at
Page copyright © 1997 Nancy B. Simmons
 Page: Tree of Life
Chiroptera. Bats.
Authored by
Nancy B. Simmons and Tenley Conway.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Chiroptera. Bats.
Authored by
Nancy B. Simmons and Tenley Conway.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Citing this page:
Simmons, Nancy B. and Tenley Conway. 1997. Chiroptera. Bats. Version 01 January 1997. http://tolweb.org/Chiroptera/15966/1997.01.01 in The Tree of Life Web Project, http://tolweb.org/