Endomychidae
Handsome fungus beetles
Floyd W. Shockley



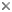
This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Endomychidae, commonly known as the Handsome Fungus Beetles, is a moderately speciose family of mycophagous beetles within the Cerylonid Series of the superfamily Cucujoidea (Crowson 1955), with 130 genera and 1782 species and subspecies (Shockley et al. 2009). Endomychidae as a family is distributed worldwide, occurring in all major biogeographical regions; however, the group is predominantly tropical with highest diversity in the Neotropics, equatorial Africa and southeast Asia. The family Endomychidae is currently arranged into 12 subfamilies (sensu Tomaszewska 2000): Anamorphinae (=Mycotheninae), Danascelinae, Endomychinae, Epipocinae, Eupsilobiinae (=Eidoreinae, Cerasommatidiidae), Leiestinae, Lycoperdininae (=Eumorphinae), Merophysiinae (=Holoparamecinae), Mycetaeinae (=Agaricophilinae), Pleganophorinae (=Trochoideinae), Stenotarsinae and Xenomycetinae.
The vast majority of taxa feed on the fruiting bodies of basidiomycete fungi, although there are representatives who specialize on softer agarics, molds, spores, and even several taxa which have been found to be facultatively phytophagous or predaceous. They rarely come to light traps and are most often collected directly from the surface of their host fungi. However, they can also be found in pitfall traps, flight intercept traps or beating/sweeping vegetation in close proximity to their hosts as they often rest on nearby foliage during the day. There are several species which have become stored product pests, and a number of taxa are inquilines of social insects, particularly from the subfamily Merophysiinae. A number of species appear to be involved in mimicry complexes with taxa from other beetle families such as Chrysomelidae, Coccinellidae, Erotylidae and Tenebrionidae, many of which are chemically defended. However, since these endomychids also tend to be chemically defended, it is unclear which is the model and which is the mimic in these Müllerian systems.
Characteristics
Adults of the family vary widely in size (ranging from 1-18 mm), morphology, and habits, particularly at the subfamilial and generic levels. Adults may be oval to elongate-oval and strongly convex to slightly flattened in habitus. Vestiture may be present or absent, but when present setae are typically recumbent to suberect. Because of this heterogeneity, it is difficult to generalize the adult morphology to make it applicable across the family. In general, the adults can be characterized by the combination of the following features (based on Lawrence et al. 1999b, Tomaszewska 2000, Skelley and Leschen 2002):
- 1-2 pairs of longitudinal sulci, sublateral lines, or subbasal foveae present (absent in some Anamorphinae, Eupsilobiinae and Mycetaeinae)
- 3-3-3 or 4-4-4 tarsal formula (may be distinctly tetramerous, pseudotrimerous, or distinctly trimerous)
- Frontoclypeal suture present
- The absence of antennal grooves (present in some Eupsilobiinae and Merophysiinae)
- Antennae generally 11-segmented (4-7 segmented in Pleganophorinae and 8-10 segmented in most Anamorphinae) with a 1-3 segmented club
- The absence of subcoxal lines on abdominal ventrite I
Similarly, the known larvae of Endomychidae are also highly variable in form, sculpture, and vestiture, and are easily confused with members of other cucujoid families. Larvae may range in size from 2-10 mm and are generally broadly ovate to elongate (fusciform in some) and strongly convex to somewhat flattened in habitus. Dorsal surfaces with vestiture of moderately long setae which may be randomly and densely distributed or sparse and aligned in transverse rows. Likewise, the vestiture may vary within the family from consisting of simple hairs to being highly specialized/modified and branched or clavate (rarely on tubercles). Endomychid larvae can be distinguished from other similar larvae by combination of the following characters (based on Lawrence 1991, Lawrence et al. 1999a):
- Head with epicranial stem short or absent and frontal arms U-shaped, V-shaped or lyriform
- 0-4 pairs of stemmata
- Antennae 3-segmented with antennomere 2 often elongate (compared to antennomeres 1 and 3)
- Mandibles unidentate to tridentate with tuberculate mola (absent in Endomychus) and usually with a well-developed prostheca (rarely reduced or absent)
- Trunk often with lateral processes on thoracic and most abdominal segments (occasionally with dorsal processes as well, and rarely these processes are deciduous)
- Legs 4-segmented with unisetose tarsungulus, urogomphi generally absent (rarely with short urogomphi on tergite IX)
Biogeography
All of the subfamilies exhibit interesting biogeographic distributional patterns. Some are cosmopolitan with genera extending over huge geographic ranges, while others are almost exclusively endemic to small, isolated regions. In general, the distributional patterns for the different subfamilies can be summarized as follows:
- Anamorphinae as a subfamily is widely distributed with genera occurring throughout the world, particularly in the tropical and subtropical regions. Species often exhibit scattered or isolated distributions, and anamorphines are often among the only endomychids found in island habitats (e.g., Caribbean Islands, Micronesian and Pacific Islands, etc.).
- Danascelinae is a small subfamily with only two genera, one of which is endemic to northern North America and the other known only from Pakistan.
- Within Endomychinae, the nominate genus Endomychus is holarctic, while the remaining genera are largely restricted to the eastern Palearctic and Indomalaysian regions.
- Epipocinae is endemic to the New World and predominantly Neotropical, though several species extend into the Nearctic region.
- Genera of Eupsilobiinae are typically endemic to small areas of Central and South America and South Africa. A notable exception is the genus Eidoreus which is cosmopolitan in distribution, but found most often in scattered island habitats.
- The subfamily Leiestinae is completely holarctic with genera restricted to either the Palearctic or the Nearctic (never both).
- The largest subfamily in terms of genera and species is Lycoperdininae. Members of this subfamily are distributed throughout the world, but most of its generic and species-level diversity is concentrated in the Indomalaysian tropics.
- Genera of the subfamily Merophysiinae are largely tropical in distribution but with restricted ranges in the Neotropical, Afrotropical and Indomalaysian regions. The genus Holoparamecus, a common stored product pest, is an exception. It has been introduced and become established in many places making it truly cosmopolitan in distribution.
- Similarly, the genus Mycetaea of the subfamily Mycetaeinae is also a stored product pest and has become established in the Nearctic, Palearctic and Afrotropical regions, while its congener Agaricophilus, which is not a pest, is endemic and restricted to eastern Europe.
- With the exception of the widespread tropical genus Trochoideus, another stored product pest, the remaining two genera of the Pleganophorinae are endemic to very small areas of the Indomalaysian and Palearctic regions, respectively.
- The subfamily Stenotarsinae is most diverse in the New and Old World tropics, though subtropical species are known from the southernmost areas of the Palearctic, the Nearctic and Australia. The largest genus in the entire family, Stenotarsus (with 250, species), is a member of this subfamily and is found throughout the tropical regions of the world (although a few species extend into the Nearctic and Palearctic regions).
- Both species of the subfamily Xenomycetinae and its constituent monotypic genus Xenomycetes are endemic to northwestern North America.
Taxonomy
Gerstaecker (1858) produced the first world monograph devoted to Endomychidae. However, most family-level treatments of the Endomychidae were limited to regional checklists and primary descriptive papers of Old or New World taxa. The consequence of this “regionalization” of the taxonomic literature is that many synonyms exist, particularly among relatively cryptic species or cosmopolitan taxa, and this problem is further compounded by insufficient global taxonomic expertise to identify those synonyms and correct the nomenclature for many of the hard-to-identify endomychid genera. Gorham (1887-1899) described many new species while reviewing the Central American fauna. Ganglbauer (1899a,1899b) reviewed the Endomychidae of Europe. In addition, several large catalogues and checklists have been produced for the family (Csiki 1901, Belon 1902, Csiki 1910, Arrow 1920, Hetschko 1926, Strohecker 1953, Rucker 2007; Tomaszewska 2007). Strohecker (1953) provided the first comprehensive keys to the known endomychid genera of the world, but this treatment lacked the phylogenetic context necessary for establishing a framework for a natural classification.
Discussion of Phylogenetic Relationships
In the 50 years since Strohecker's work (1953), very little work has been done to investigate the phylogenetic relationships of the taxa contained within the family, as well as the relationship of Endomychidae to other cucujoid families. Inclusion of the Eupsilobiinae and the Anamorphinae in Endomychidae has led some researchers to suggest that the family may in fact be polyphyletic (Slipinski & Pakaluk 1991, Robertson et al. 2008), particularly with respect to the families Coccinellidae and Corylophidae. Historically, the Endomychidae were considered to be the sister taxon to the more well-known, predominantly predaceous Coccinellidae. In fact, some older classifications placed these two taxa together under the name "Trimera", based on the shared feature of pseudotrimerous tarsi (Tomaszewska 2000). Of course, this condition within Endomychidae is now known to be restricted to the “higher” endomychid subfamilies—Endomychinae, Epipocinae, Lycoperdininae and Stenotarsinae (Tomaszewska 2005).
Because of Endomychidae’s potential sister-group association with the economically significant Coccinellidae, much interest has arisen in recent years in performing phylogenetic analyses to investigate these relationships more thoroughly. Tomaszewska (2000) performed the first formal phylogenetic analysis of the family based on morphological characters of the adults; however, the results only supported the monophyly of the family and offered no resolution between the subfamilies. In a follow-up paper that included larval characters, Tomaszewska (2005) was able to obtain better resolution. Unfortunately, both studies assumed a sister-group relationship with Coccinellidae rather than including sufficient outgroup taxa sampled from throughout the Cerylonid-Series and the superfamily Cucujoidea.
Hunt et al. (2007), while presenting a phylogeny for the entire order Coleoptera based on 3 genes (18S, 16S, COI) recovered a monophyletic Cerylonid-Series with the same “higher” Endomychidae sister group relationship with Coccinellidae, but recovered the Merophysiinae, Leiestinae and Anamorphinae clustered together and paraphyletic with respect to Corylophidae. Of course, the monophyly of Endomychidae was not the focus of that study, nor was its taxon or gene sampling sufficient to reliably resolve the familial and subfamilial relationships of Endomychidae and the other Cerylonid-Series families. Nonetheless, its resulting hypotheses for the placement of some of the endomychid subfamilies raised some interesting and lingering questions about the monophyly of the Endomychidae which warranted further exploration.
Robertson et al. (2008) conducted the first formal test of the limits of the Cerylonid-Series using molecular sequence data (18S, 28S). The conclusion of that study was that the monophyly of the family Endomychidae was not supported as it is currently defined. While they did recover a monophyletic clade of subfamilies corresponding to the “higher” Endomychidae of Tomaszewska sister to Coccinellidae, Robertson et al. (2008) recovered Anamorphinae outside of Endomychidae and sister to Corylophidae. Anamorphinae being recognized as an independent assemblage outside of the remaining Endomychidae was first proposed by Sasaji (1978, 1987, 1990) as Mychothenidae, but this classification was never widely accepted or implemented. Clearly, much work is needed to fully resolve the family and its constituent subfamilies. For the purposes of the tree presented above, I have opted to use the phylogenetic hypothesis proposed by Tomaszewska (2005), despite its outgroup limitations. At present, that analysis remains as the most robust phylogenetic analysis of Endomychidae to include members of all 12 subfamilies.
References
Arrow, G. J. 1920. A contribution to the classification of the coleopterous family Endomychidae. Transactions of the Entomological Society of London 1920: 1-83.
Belon, R. P. 1902. Coleoptera Clavicornia Fam. Lathridiidae, pp. 1-40 + 1 pl. In P. Wytsman [ed.], Genera Insectorum. P. Wytsman, Brussels.
Crowson, R.A. 1955. The Natural Classification of the Families of Coleoptera. Lloyd, London. 187 pp.
Csiki, E. 1901. Catalogus Endomychidarum. Termeszetrajzi Fuzetek 24: 2-53.
Csiki, E. 1910. Pars 12: Fam. Endomychidae, pp. 1-68. In S. Schenkling [ed.], Coleopterorum Catalogus. W. Junk, Berlin.
Ganglbauer, L. 1899a. Familienreihe Staphylinoidea. II. Theil. & Familienreihe Clavicornia. XXIX. Familie. Lathridiidae, pp. 766-821. Carl Gerolds Sohn, Wien.
Ganglbauer, L. 1899b. Familienreihe Staphylinoidea. II. Theil. & Familienreihe Clavicornia. XXXII. Familie. Endomychidae, pp. 909-941. Die Kafer von Mitteleuropa. Carl Gerolds Sohn, Wien.
Gerstaecker, A. 1858. Monographie der Endomychidae, einer Familie der Coleopteren, pp. XIV + 433 pp., 3 pls. In A. Gerstaecker [ed.], Entomographien. Abhandlungen in Bereich der Gliederthiere, mit besonderer Benutzung der Koenigl. Entomologischen Sammlung zu Berlin. W. Engelmann, Leipzig.
Gorham, H. S. 1887-1899. Biologia Centrali-Americana. Insecta. Coleoptera. Vol. VII. Erotylidae, Endomychidae, and Coccinellidae. VII. London. 276 pp., 13 lams pp.
Hetschko, A. 1926. Pars 85: Lathridiidae, pp. 1-86. In S. Schenkling [ed.], Coleopterorum Catalogus. W. Junk, Berlin.
Hunt, T., J. Bergsten, Z. Levkanicova, A. Papadopoulou, O. St. John, R. Wild, P. M. Hammond, D. Ahrens, M. Balke, M. S. Caterino, J. Gomez-Zurita, I. Ribera, T. G. Barraclough, M. Bocakova, L. Bocak, and A. P. Vogler. 2007. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 318: 1913-1916.
Lawrence, J. F. 1991. Endomychidae (Cucujoidea)(including Merophysiidae, Mycetaeidae), pp. 482-485. In F. W. Stehr [ed.], Immature Insects. Volume 2. Kendall/Hunt Publishing Company, Dubuque, IA.
Lawrence, J. F., and A. F. Newton. 1995. Families and subfamilies of Coleoptera (with selected genera, notes, references and data on family-group names), pp. 779-1006. In J. Pakaluk and S. A. Slipinski [eds.], Biology, Phylogeny, and Classification of Coleoptera: Papers Celebrating the 80th Birthday of Roy A. Crowson. Museum i Instytut Zoologii PAN, Warszawa.
Lawrence, J. F., A. M. Hastings, M. J. Dallwitz, T. A. Paine, and E. J. Zurcher 1999a. Endomychidae. Beetle Larvae of the World: Descriptions, Illustrations, Identification, and Information Retrieval for Families and Sub-families. CD-ROM, version 1.1 for MS-Windows. CSIRO Publishing, Melbourne.
Lawrence, J. F., A. M. Hastings, M. J. Dallwitz, T. A. Paine, and E. J. Zurcher 1999b. Endomychidae (major part). Beetles of the World: A Key and Information System for Families and Subfamilies. CD-ROM, version 1.0 for MS-Windows. CSIRO Publishing, Melbourne.
Robertson, J. A., M. F. Whiting, and J. V. McHugh. 2008. Searching for natural lineages within the Cerylonid Series (Coleoptera: Cucujoidea). Molecular Phylogenetics and Evolution 46: 193-205.
Rucker, H.W. and I. Lobl. 2007. Endomychidae (subfamily Merophysiinae), pp. 557-559. In: I. Lobl and A. Smetana (eds.), Catalogue of Palaearctic Coleoptera, Vol. 4. Apollo Books, Stenstrup. 935 pp.
Sasaji, H. 1978. Notes on the Japanese Endomychidae, with an establishment of a new subfamily (Coleoptera). Memoirs of the Faculty of Education, Fukui University Series II (Natural Science) 28: 1-31.
Sasaji, H. 1987. On the higher classification of the Endomychidae and their relative families (Coleoptera). Entomological Journal of Fukui 1: 44-51.
Sasaji, H. 1990. The family Mychothenidae of Japan (Coleoptera). Esakia Special Issue No. 1: 65-75.
Shockley, F.W., K.W. Tomaszewska and J.V. McHugh. 2009. An annotated checklist of the handsome fungus beetles of the world (Coleoptera: Cucujoidea: Endomychidae). Zootaxa 1999: 1-113.
Skelley, P.E. and R.A.B. Leschen. 2002. Endomychidae. Pp. 366-370. In: R. H. Arnett, Jr., M.C. Thomas, P.E. Skelley, and J.H. Frank, eds. American Beetles. Vol. 2. Polyphaga: Scarabaeoidea through Curculionidae. CRC Press LLC, Boca Raton, xiv + 861 pp.
Slipinski, S.A. and J. Pakaluk. 1991. Problems in the classification of the Cerylonid series of Cucujoidea (Coleoptera). Pp. 79-88. In: M. Zunino, X. Belles and M. Blas, eds. Advances in Coleopterology. European Association of Coleopterology. Silvestrelli and Cappelletto. Torino.
Strohecker, H.F. 1953. Coleoptera. Fam. Endomychidae. In: P. Wytsman, ed. Genera Insectorum. Fasc. 210. L. Desmet-Verteneuil, Brussels, 140 pp.
Strohecker, H.F. 1986. A Catalog of the Coleoptera of America North of Mexico. Family: Endomychidae. United States Department of Agriculture (Agriculture Handbook 529-98), Washington, DC. viii + 19 pp.
Tomaszewska, K.W. 2000. Morphology, phylogeny and classification of adult Endomychidae (Coleoptera: Cucujoidea). Annales Zoologici 50 (4): 449-558.
Tomaszewska, K. W. 2005. Phylogeny and generic classification of the subfamily Lycoperdininae with a re-analysis of the family Endomychidae (Coleoptera: Cucujoidea). Annales Zoologici (Warszawa) Supplement: 1-172.
Tomaszewska, W.K. 2007. Endomychidae (remaining subfamilies), pp. 559-568. In: I. Lobl and A. Smetana (eds.), Catalogue of Palaearctic Coleoptera, Vol. 4. Apollo Books, Stenstrup. 935 pp.
Title Illustrations

| Scientific Name | Endomychidae |
|---|---|
| Specimen Condition | Live Specimen |
| Life Cycle Stage | Adult |
| View | dorsal |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© Floyd W. Shockley

|
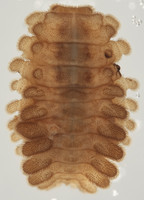
| Scientific Name | Anidrytus sp. |
|---|---|
| Specimen Condition | Dead Specimen |
| Identified By | Floyd W. Shockley |
| Life Cycle Stage | Larva |
| View | dorsal |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© Floyd W. Shockley
|

| Scientific Name | Anamorphus sp. |
|---|---|
| Specimen Condition | SEM |
| Identified By | Floyd W. Shockley |
| Life Cycle Stage | Adult |
| View | dorsal |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© Floyd W. Shockley
|
About This Page
Special thanks to Joseph V. McHugh for reviewing the text and offering useful comments/suggestions. Partial support for the construction of this page was provided by the H.H. Ross Endowment of the Department of Entomology at the University of Georgia, through an NSF AToL grant EF-0531665 to M.F. Whiting (subcontract to J.V. McHugh) and through an NSF PEET grant (DEB-0329115) to J.V. McHugh, M.F. Whiting, and K.B. Miller.
Floyd W. Shockley
Dept. of Entomology, National Museum of Natural History, Smithsonian Institution
Correspondence regarding this page should be directed to Floyd W. Shockley at
Page copyright © 2011 Floyd W. Shockley
 Page: Tree of Life
Endomychidae. Handsome fungus beetles.
Authored by
Floyd W. Shockley.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Endomychidae. Handsome fungus beetles.
Authored by
Floyd W. Shockley.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 23 June 2008
- Content changed 31 March 2009
Citing this page:
Shockley, Floyd W. 2009. Endomychidae. Handsome fungus beetles. Version 31 March 2009. http://tolweb.org/Endomychidae/9169/2009.03.31 in The Tree of Life Web Project, http://tolweb.org/